Il tessuto muscolare è caratterizzato dalla sua peculiare capacità di contrazione che permette di garantire i movimenti delle strutture anatomiche di cui fa parte.
Il tessuto muscolare è formato da fasci di cellule organizzate in modo da conferire al tessuto una peculiare capacità di contrazione. Nell’organismo umano si distinguono tre tipi di tessuti muscolari:
– il tessuto muscolare scheletrico;
– il tessuto muscolare cardiaco;
– il tessuto muscolare liscio
(Fig. 1).
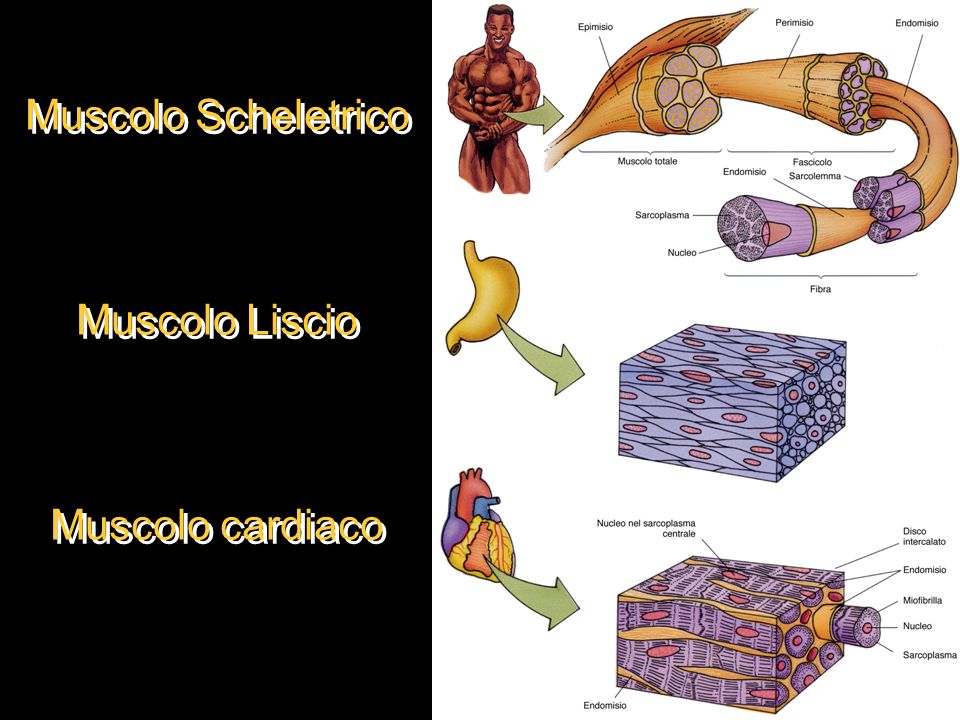
Figura 1: I tre tipi di tessuto muscolare del corpo umano
Tessuto muscolare scheletrico
Il tessuto muscolare striato scheletrico forma i muscoli dell’apparato locomotore, ma è presente anche in alcuni organi dell’apparato digerente e respiratorio. Esso è deputato principalmente a produrre movimento, ma ha anche la funzione di mantenere la postura, proteggere gli organi interni e mantenere la temperatura corporea.
Gli elementi che lo compongono sono le fibre muscolari, ossia cilindri lunghi da 1 mm a 20 cm, formate da numerose cellule i cui nuclei sono localizzati a livello della membrana plasmatica o sarcolemma.
Ogni fibra è rivestita da tessuto connettivo fibrillare lasso detto endomisio; l’insieme di più fibre forma un fascio muscolare, rivestito anch’esso da connettivo lasso che forma il perimisio; infine l’insieme dei fasci forma il muscolo, rivestito da una lamina connettivale più densa, l’epimisio (Fig. 2).
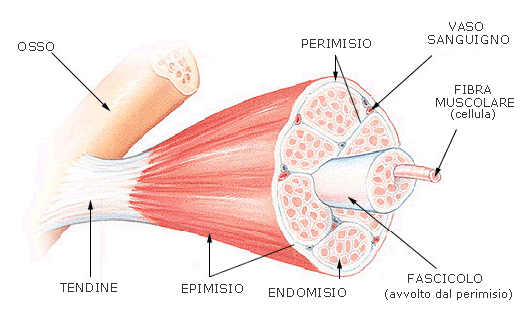
Figura 2: organizzazione strutturale del muscolo striato scheletrico
Il sarcoplasma delle fibre muscolari mostra una striatura trasversale caratterizzata dall’alternanza di bande chiare (banda I) e bande scure (bande A). Al centro di ogni banda chiara è presente una stria scura (linea Z), mentre al centro di ogni banda scura vi è una stria chiara (linea H) con al centro un’ulteriore stria scura sottile (linea M).
Lo spazio interposto tra due linee Z definisce il sarcomero, ovvero l’unità funzionale della fibra muscolare. Il sarcomero è formato da un insieme di due tipi di miofilamenti, spessi e sottili. I filamenti sottili sono composti in prevalenza dalla proteina actina, mentre i filamenti spessi sono costituiti da miosina. Le zone in cui si verifica sovrapposizione di miosina e actina corrispondono alle bande scure, mentre nelle bande chiare si ritrova una sola delle due proteine (Fig. 3).
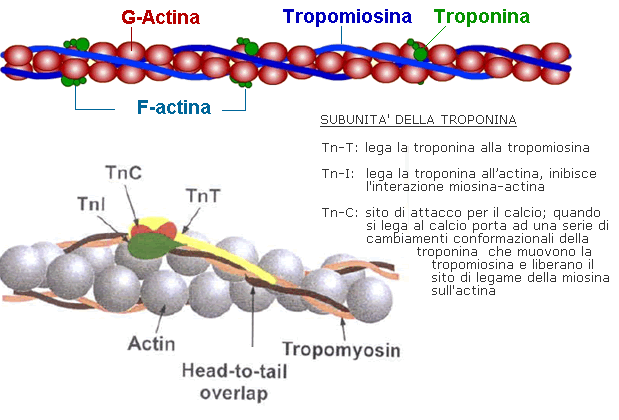
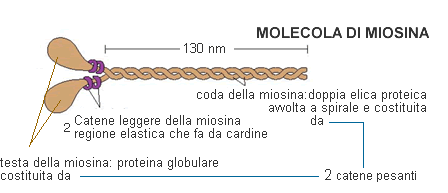
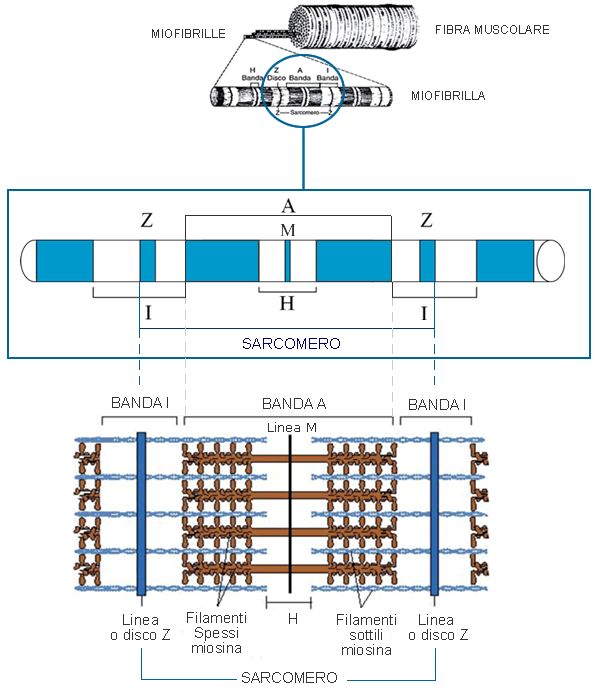
Figura 3: Rappresentazione della struttura dei filamenti spessi di miosina, dei filamenti sottili di actina- bande scure e chiare
L’actina e la miosina rappresentano le principali proteine contrattili del muscolo scheletrico, insieme alla tropomiosina e alla troponina.
- La miosina è la proteina più abbondante, è formata da una coda e una testa, dotata di attività ATPasica. Ogni miofilamento spesso è formato da un fascio di numerose molecole di miosina le cui teste sporgono dal filamento stesso;
- Le molecole di actina sono globulari (G-actina) e in ogni miofilamento sottile sono legate tra loro a formare lunghe file avvolte ad elica (F-actina);
- La tropomiosina è formata da due catene avvolte a spirale che vanno ad inserirsi nelle catene di F-actina;
- La troponina è costituita da tre subunità globulari: troponina I deputata all’inibizione dei contatti actina-miosina, troponina C che lega gli ioni calcio e troponina T che media il contatto con la tropomiosina.La contrazione avviene per scivolamento dei filamenti sottili lungo i filamenti spessi.In condizioni di riposo le molecole di troponina impediscono alla miosina di legarsi all’actina. Quando arriva l’impulso nervoso per la contrazione, si ha il rilascio di acetilcolina, un neurotrasmettitore che causa la depolarizzazione del sarcolemma. Ciò porta al rilascio di ioni calcio, che legandosi alla troponina ne cambiano la conformazione molecolare liberando la miosina e attivando la funzione ATPasica delle teste di miosina. L’attivazione di tale funzione libera l’energia necessaria per il legame delle teste di miosina all’actina. I prodotti dell’idrolisi dell’ATP si distaccano dalla testa della miosina e una nuova molecola di ATP si lega ad essa; in tal modo avviene il distacco tra la miosina e l’actina e il ciclo può ricominciare. Se una nuova molecola di ATP non si lega alla miosina si verifica rigidità muscolare. Questi eventi avvengono contemporaneamente in tutti i sarcomeri successivi della fibra muscolare e allo stesso tempo in tutte le miofibrille della fibra, pertanto quest’ultima subisce un accorciamento (contrazione).
La contrazione del tessuto muscolare scheletrico è controllata dal sistema nervoso centrale ed avviene in seguito a stimoli derivati dalle fibre nervose del SNC su aree specializzate dette placche motrici.I nervi penetrano nell’epimisio, si ramificano attraverso il perimisio e penetrano nell’endomisio per innervare le singole fibre muscolari. La comunicazione chimica tra il terminale sinaptico di un neurone e la placca motrice della fibra muscolare scheletrica avviene a livello della giunzione neuromuscolare (Fig. 4).In ogni muscolo ciascuna fibra muscolare è innervata da un solo motoneurone ma ogni motoneurone innerva più fibre muscolari. L’insieme delle fibre muscolari controllate da un singolo motoneurone costituisce un’unità motoria. Alcune unità motorie rimangono attive anche quando il muscolo è a riposo; le loro contrazioni risultano insufficienti per provocare uno spostamento e servono a mantenere il muscolo in uno stato di tensione definito tono muscolare.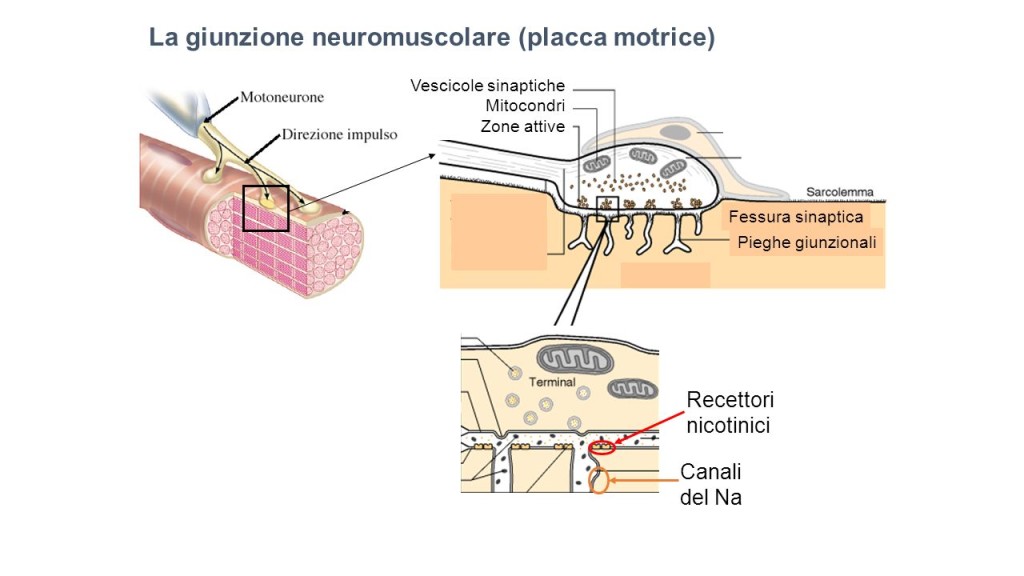
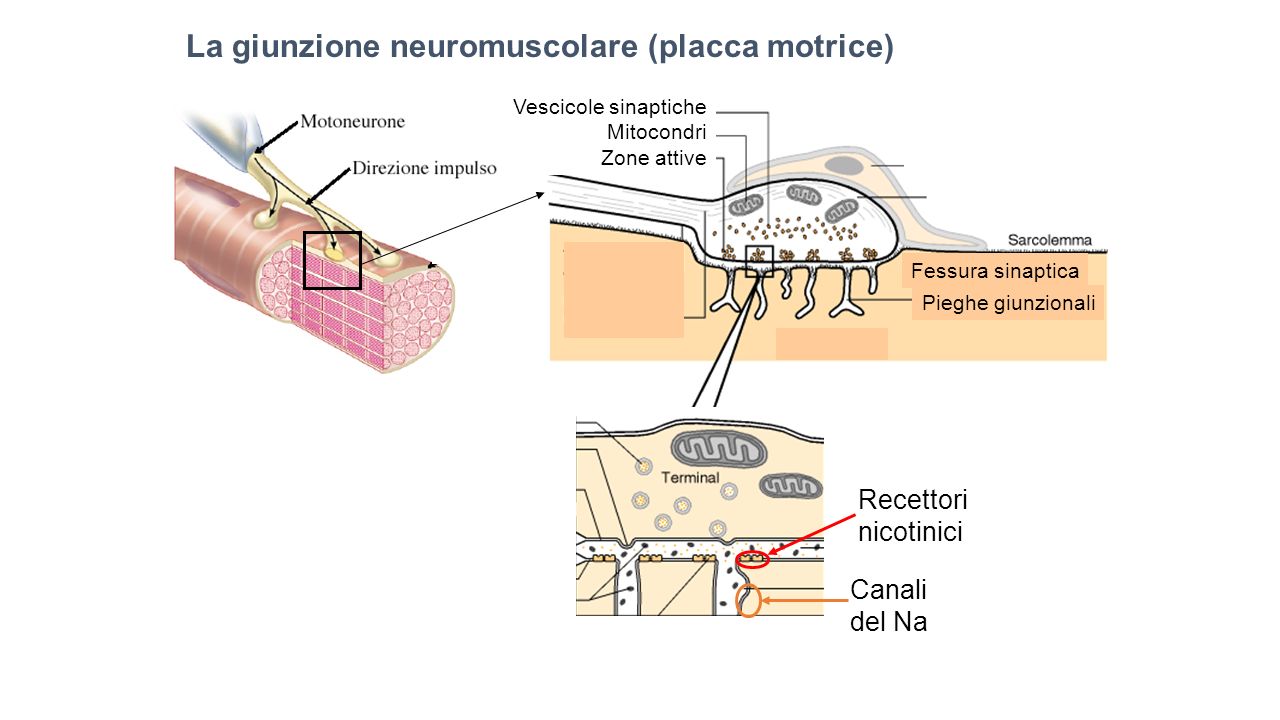
Figura 4: Rappresentazione schematica della giunzione neuromuscolare
Tessuto muscolare liscio
Il tessuto muscolare liscio è formato da cellule muscolari lisce, ossia cellule di forma allungata e fusata, provviste di miofilamenti privi di striature. Tali cellule per contrarsi sfruttano ugualmente i filamenti di actina e miosina che, a differenza del tessuto scheletrico, non sono organizzati in miofibrille. La contrazione delle cellule muscolari lisce è lenta ma può essere prolungata nel tempo con un basso consumo di energia rispetto al tessuto muscolare striato. Anche in questo caso la contrazione avviene per scorrimento reciproco dei filamenti di actina su quelli di miosina, ma la proteina coinvolta nell’attivazione della contrazione è la calmodulina. L’interazione calcio- calmodulina attiva la chinasi della catena leggera della miosina che fosforila la miosina permettendone il legame con l’actina. Gli stimoli per la contrazione possono essere di origine nervosa, ormonale o farmacologica.
Le cellule muscolari lisce generalmente, si organizzano a formare fasci nella parete degli organi cavi, compresi i vasi sanguigni, nella pelle e nel bulbo oculare.
Tessuto muscolare cardiaco
Il tessuto muscolare striato cardiaco è costituito da singole fibrocellule, o cardiomiociti, collegate tra loro mediante giunzioni intercellulari, dette dischi intercalari. Pertanto da un punto di vista funzionale il muscolo cardiaco si comporta come un sincizio pur non essendolo da un punto di vista strutturale. I cardiomiociti hanno un singolo nucleo situato al centro e si presentano ramificati alle estremità; possiedono inoltre i sarcomeri e le caratteristiche striature trasversali tipiche delle fibre muscolari scheletriche. La contrazione muscolare avviene con le stesse modalità del tessuto muscolare scheletrico e coinvolge i cardiomiociti del cosiddetto miocardio comune, che costituisce la gran parte del tessuto muscolare del cuore. L’altra componente è il miocardio specifico che ha la funzione di condurre ad alta velocità l’impulso di contrazione (Fig. 6).
Il tessuto miocardico si contrae involontariamente e il suo ritmo è controllato dal sistema nervoso autonomo.

Figura 6: Organizzazione strutturale del tessuto muscolare striato cardiaco.